THE HELIX
 The helix links to biotensegrity through a common origin in the geodesic geometry of the platonic solids (see geodesic and helix pages). Helical molecules behave as tensegrity structures in their own right as they naturally stabilize through a balance between attraction (tension) and repulsion (compression).Connelly; Edwards Globular proteins contain multiple helical domains and can polymerize into larger helixes such as those in the cytoskeleton described above. Similar helices can form into heterarchies as they wind around each other to form coiled-coils (eg. spectrin) or assemble into mechanically rigid rods or filaments,Luo; Qin or further combine into more complex structures with specialized functions (eg. collagens). Collagens are major structural proteins that consist of several heterarchical levels of helices, and are the main structural protein within bones, tendons, ligaments and fascia.
The helix links to biotensegrity through a common origin in the geodesic geometry of the platonic solids (see geodesic and helix pages). Helical molecules behave as tensegrity structures in their own right as they naturally stabilize through a balance between attraction (tension) and repulsion (compression).Connelly; Edwards Globular proteins contain multiple helical domains and can polymerize into larger helixes such as those in the cytoskeleton described above. Similar helices can form into heterarchies as they wind around each other to form coiled-coils (eg. spectrin) or assemble into mechanically rigid rods or filaments,Luo; Qin or further combine into more complex structures with specialized functions (eg. collagens). Collagens are major structural proteins that consist of several heterarchical levels of helices, and are the main structural protein within bones, tendons, ligaments and fascia.
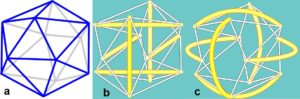
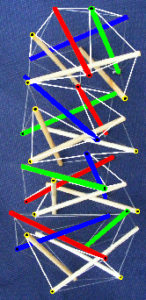 Axial stretching or compression of a simple tensegrity helix initiates rotation in a direction that depends on the direction of twist or chirality. Inter linking it to another one with opposite chirality causes resistance as each helix counteracts the rotation of the other. Crossed-fibres of collagen scale up to form tubular helixes in the walls of blood vessels, the urinary system and intestinal tractGabella and influence their mechanical properties. Elastic arteries such as the aorta have walls organized into lamellar units, with collagen reinforcement and smooth muscle cells that form crossed-helices. Tension helical ‘walls’ will inevitably contain and interact with other structures under compression, and at different heterarchical levels. This chain of T-icosa shows how the (coloured) struts form both left- and right-handed helices, with each part remaining distinct, just like in these higher-level examples.
Axial stretching or compression of a simple tensegrity helix initiates rotation in a direction that depends on the direction of twist or chirality. Inter linking it to another one with opposite chirality causes resistance as each helix counteracts the rotation of the other. Crossed-fibres of collagen scale up to form tubular helixes in the walls of blood vessels, the urinary system and intestinal tractGabella and influence their mechanical properties. Elastic arteries such as the aorta have walls organized into lamellar units, with collagen reinforcement and smooth muscle cells that form crossed-helices. Tension helical ‘walls’ will inevitably contain and interact with other structures under compression, and at different heterarchical levels. This chain of T-icosa shows how the (coloured) struts form both left- and right-handed helices, with each part remaining distinct, just like in these higher-level examples.
Having said all this, one of the characteristics of the tensegrity-icosahedron model (T-icosa) is that it consists of 3 struts arranged in a left-handed twist and 3 struts with a right-handed twist, and when opposite sides are pushed together, the entire structure contracts symmetrically around a central point and decreases in volume. In other words, tensegrities can demonstrate auxetic behaviour that was once considered impossible, and has now been shown in tendons, skin, bone and organs etc. So, some of the standard mechanical descriptions of helices given here may need some tweaking when we add this auxetic property, particularly in the way muscles function, but this is ongoing at present.
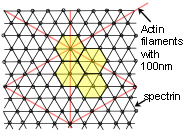
Capillary formation results from tension-dependent interactions between endothelial cells and an extra-cellular scaffold of their own construction and these cells form a selective barrier that allows vascular contents to pass out between capillary walls. The internal cellular cytoskeleton determines cell shape and orientation through tensegrity, is affected by signalling mechanisms and variations in fluid flow, and alters the tension between cells through adherens junctions, and thus ultimately affects tube permeability. This could perhaps be compared with the wall of a helical tensegrity model that has many gaps, but if the struts could be expanded into plates that just touched each other so that they ‘sealed’ the internal space; it could act just like the capillary cells.
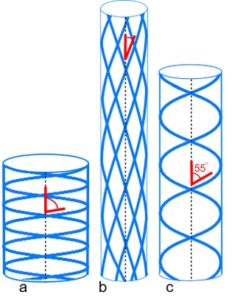
 In 1956, it was definitively shown that an optimum helical angle of ~55o (relative to the main helical axis) balances both longitudinal and circumferential stresses,Clark; Shadwick and thus allow pressurized tubes to bend smoothly without kinking and resist torsional deformation. Cardiac muscle fibre orientation varies linearly between inner and outer walls, from 55o in one direction to 55o in the other, with tangential spiralling in a transverse plane. The heart is a helical coil of muscle that contracts with left and right-handed twisting motions,buckberg and has been described Stephen Levin as a simple tensegrity pump that may have relevance to cardiac dynamics, and has also been described using the ‘jitterbug’ mechanism (see Geodesic page).Levin; Verheyen; Judge
In 1956, it was definitively shown that an optimum helical angle of ~55o (relative to the main helical axis) balances both longitudinal and circumferential stresses,Clark; Shadwick and thus allow pressurized tubes to bend smoothly without kinking and resist torsional deformation. Cardiac muscle fibre orientation varies linearly between inner and outer walls, from 55o in one direction to 55o in the other, with tangential spiralling in a transverse plane. The heart is a helical coil of muscle that contracts with left and right-handed twisting motions,buckberg and has been described Stephen Levin as a simple tensegrity pump that may have relevance to cardiac dynamics, and has also been described using the ‘jitterbug’ mechanism (see Geodesic page).Levin; Verheyen; Judge
Similar helices form heterarchical ‘tubes within tubes’ in fascia that permeate and surround the muscles, limbs and body walls of a huge variety of species, all considered through biotensegrity (see myofascial helices page). Tubular organs that maintain constant volume throughout changes in shape have been described in the tongues of mammals and lizards, the arms and tentacles of cephalopods and the trunks of elephants.Kier
The arrangement of scales on the pangolin illustrate the helix at the macro level, although notice how the orientations of left and right-handed helices on the body are different in the limbs, and this pattern in the limbs may be related to the Fibonacci sequence (see Palpatory phenomena and Traube-Hering-Mayer waves page). The thoraco-lumbar and abdominal fasciae also have a spiral appearance, if only in part, and helical fascial sheaths that transfer tensional forces within and between themselves have been described in controlling movement in a way that the nervous system is incapable of.Stecco and Myers
COLLAGEN
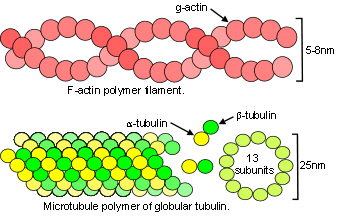
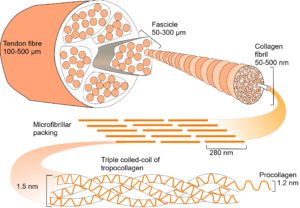 Bones, tendons, ligaments and fascia are all arranged into complex heterarchy/hierarchies with collagen appearing at several different levels within them. In collagen type I, repeating sequences of amino acids spontaneously form a left-handed helix of procollagen, with three of these combining to form a right-handed tropocollagen molecule. Five tropocollagen molecules then coil in a staggered helical array that lengthens longitudinally by the addition of more tropocollagen to form a microfibril, and pack radially to form a fibril;Charvolin with higher arrangements forming fibres and then fascicles. (see helix page).
Bones, tendons, ligaments and fascia are all arranged into complex heterarchy/hierarchies with collagen appearing at several different levels within them. In collagen type I, repeating sequences of amino acids spontaneously form a left-handed helix of procollagen, with three of these combining to form a right-handed tropocollagen molecule. Five tropocollagen molecules then coil in a staggered helical array that lengthens longitudinally by the addition of more tropocollagen to form a microfibril, and pack radially to form a fibril;Charvolin with higher arrangements forming fibres and then fascicles. (see helix page).
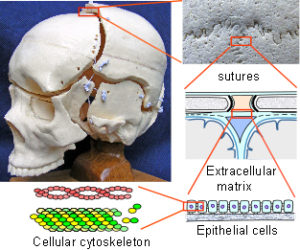
The collagen molecule exists in twenty-eight different types, or configurations, and is a major structural contributor to the extracellular matrix (ECM) that surrounds virtually every cell. The matrix attaches to the cellular cytoskeleton through adhesion molecules in the cell membrane and forms a structural framework that extends through the fascia to every level in the body.
FASCIAL SYSTEM
Traditionally considered as mere packing tissue, fascia has been show to exert considerable influence over muscle generated force transmission.Huijing; Stecco Fascial architecture consists of a complex network of tensioned fibres that envelop and enclose a heterarchical system of compressed compartments at multiple size-scales, and that are particularly noticeable in cross-section of the limbs as ‘tubes within tubes’. At the higher scale, these compartments contain muscles, bones and organs, while at the meso scale, the compartments are continuous with blood vessels, nerves and other tubes; with cells and proteoglycan-bound fluids of the extracellular matrix occupying the micro scale compartments; all of which operate as parts of a whole.
Within muscle a delicate network of endomysium surrounds individual muscle cells and is continuous with the perimysium ensheathing groups of fibres in parallel bundles, or fascicular compartments. Perimysial septa are themselves inward extensions of the epimysium, which covers the muscle and is continuous with the fascia investing whole muscle groups. These fascial tissues are reinforced by two helical crossed-ply sets of collagen with the ‘ideal’ resting fibre orientation of 55o (axial) that varies with changing muscle length (see myofascial helices page).
Tension and compression are functionally interacting with each other, and because they always occur together, this combination of tensioned fibres and compressed compartments is what gives the body its form and the fascia its functions.
Over the last few years, the surgeon Jean-Claude Guimberteau 1 and 2 has observed and described the fibrous extracellular matrix system in vivo that forms a histological continuum between bones, tendons fascia, skin, muscles and vasculature etc, and allows them to glide in relation to each other during movement. The tensioned fibres form polyhedral ‘microvacuoles’ that contain fluid-bound proteoglycans and form the most basic level of tissue organization, where (at the microscopic level) there is no clear distinction between where one structure ends and another begins. The fibres are, in effect, the bars of a closed kinematic chain (CKC) system that enables each one to guide and regulate the position and motion of all the others within the system. CKCs are ubiquitous in biology and form the basic mechanics of the tensegrity model – they are described more fully here and in the book.
The following now gives some examples of how biotensegrity changes the way that we think about the human body.
THE HUMAN CRANIUM
 Many aspects of normal cranial development are poorly understood, with many of the old ideas now outdated, and the biotensegrity model explains some of these and has the potential to improve our understanding of normal and abnormal cranial development. A more detailed explanation is given on the cranial vault page.
Many aspects of normal cranial development are poorly understood, with many of the old ideas now outdated, and the biotensegrity model explains some of these and has the potential to improve our understanding of normal and abnormal cranial development. A more detailed explanation is given on the cranial vault page.
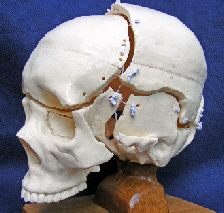
The skull is generally considered to be a solid box but is actually made up of 22 bones, most of which remain distinct throughout life, and several of these contribute to the cranial vault that surrounds and protects the brain. The sutural spaces between the bones are filled with fibrous tissue and are important to the mechanism that allows the cranium to grow larger and accommodate the developing brain. A tough dural membrane (dura mater) lines the internal surface of the bones and passes down through the cranial cavity to the base of the skull. Until recently the general opinion was that the growing brain pushes the bones outwards but this is now known not to be the case; and although an increase in dural tension does stimulate bone growth, the mechanism is much more complex than previously thought and is now better explained through biotensegrity.
 In this example, the geodesic dome (icosahedron) is developed into a T-icosa model with the struts connecting opposite vertices (almost). The straight struts are then replaced with curved struts and these are replaced with curved plates (not shown) to produce the model skull with bones that surround a central space. The bones of the cranial vault are tensioned by the dura mater and configured as a tensegrity structure. Note that curved struts only make sense in biology because they are at the top of complex structural heterarchies (at least nine different levels within bone) and that extend from the molecular level to the complete structure at the macro level (see definitions page). It should also be recognized that the vault is also influenced by the outer muscles and fascia as well as the cerebral hemispheres and internal tissues.
In this example, the geodesic dome (icosahedron) is developed into a T-icosa model with the struts connecting opposite vertices (almost). The straight struts are then replaced with curved struts and these are replaced with curved plates (not shown) to produce the model skull with bones that surround a central space. The bones of the cranial vault are tensioned by the dura mater and configured as a tensegrity structure. Note that curved struts only make sense in biology because they are at the top of complex structural heterarchies (at least nine different levels within bone) and that extend from the molecular level to the complete structure at the macro level (see definitions page). It should also be recognized that the vault is also influenced by the outer muscles and fascia as well as the cerebral hemispheres and internal tissues.
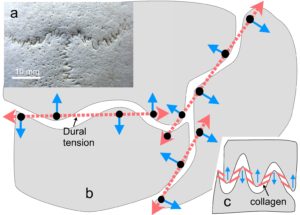 Adult bones are separated by a sutural gap of about 100 microns and have large curves and smaller serrated outlines with a fractal relationship between them.Yu The interaction between the collagen fibres within the dural membrane and bone convexities causes the bones to be held apart with the alignment of collagen fibres in the serrated sutures acting similarly,Jasinoski and both contribute to the tensegrity configuration. (see definitions page).
Adult bones are separated by a sutural gap of about 100 microns and have large curves and smaller serrated outlines with a fractal relationship between them.Yu The interaction between the collagen fibres within the dural membrane and bone convexities causes the bones to be held apart with the alignment of collagen fibres in the serrated sutures acting similarly,Jasinoski and both contribute to the tensegrity configuration. (see definitions page).
During growth, the vault bones develop totally within the membrane, which they separate into an outer periosteum and inner dura mater membrane as they grow around their edges (bone fronts). Tension in the dural membrane beneath the sutures, plus chemical signals from the osteoblasts (bone-making cells) at the bone fronts, influence the cytoskeleton of cells within the membrane and beneath the suture through the process of mechanotransduction, and thus change cell activity that increases bone growth. It is a cyclic mechanism that both regulates bone development and maintains sutural patency up until the age of about seven years (when the brain stops growing). Even after this age the sutures normally remain patent up until at least the age of 70 years and contribute to the small amounts of bone mobility recognized by ‘cranial’ osteopaths and ‘cranio-sacral’ therapists (see Palpatory phenomena and Traube-Hering-Mayer pages).
 The bones form a dome that provides protection to the brain, compression struts of a tensegrity architecture that maintains sutural flexibility and accommodates brain growth, and a microstructure that transfers external forces down through through the heterarchy to the nano-scale. The centre of the bone is a honeycomb like structure made from collagen and mineral reinforcement. Curved-strut plates are still compatible with biotensegrity when considered in terms of heterarchies because the forces of tension and compression are always acting in straight lines at some smaller scale.
The bones form a dome that provides protection to the brain, compression struts of a tensegrity architecture that maintains sutural flexibility and accommodates brain growth, and a microstructure that transfers external forces down through through the heterarchy to the nano-scale. The centre of the bone is a honeycomb like structure made from collagen and mineral reinforcement. Curved-strut plates are still compatible with biotensegrity when considered in terms of heterarchies because the forces of tension and compression are always acting in straight lines at some smaller scale.
A biotensegrity configuration allows the skull to enlarge and remain one step in front of the growing brain rather than being physically pushed out by it. It also allows the skull to respond to the mechanical demands of external muscular and fascial structures and integrate the entire cranium. The dural membrane that lines the inner surface of the bones also extends into the cranial cavity; and it may be that ‘aberrant’ stresses in the base of the newborn skull are transferred to the bones of the vault (through the dura) and alter their normal growth pattern, thus leading to distortions in head shape (plagiocephaly) and sutural fusion (craniosynostosis).
SPIDER SILK
Spider silk can also be considered as a tensegrity structure at bothe the microKnight and macro levels.Connelly It is a composite material with a heterarchical structure composed mainly of the proteins Spidroin I and II. Spidroin I consists of poly-alanine chains in anti-parallel beta-sheet conformation packed into an orthorhombic crystallite unit. These crystallites are interconnected by helical oligopeptides rich in glycine that form a polypeptide chain network within an amorphous glycine-rich matrix. The overall network shape contains circular segments (40-80 nm diameters) interconnecting in series to form a silk fibril, with many of these arranged laterally to form the silk thread with a diameter of 4-5 microns.Termonia; Du It is the regular spacing and orientation of these crystallite units and heterarchical structure that suggests that it is a tensegrity structure.

 An analogy can also be made between a spider’s web and the spoked wheel where cable tension is balanced by compression within the rim and central hub. If the cables were fairly flexible the central hub could be moved somewhat and always return to the same position of stable equilibrium – an important and most-efficient tensegrity characteristic called super-stabilityConnelly.
An analogy can also be made between a spider’s web and the spoked wheel where cable tension is balanced by compression within the rim and central hub. If the cables were fairly flexible the central hub could be moved somewhat and always return to the same position of stable equilibrium – an important and most-efficient tensegrity characteristic called super-stabilityConnelly.
THE BICYCLE WHEEL AND SHOULDER JOINT
 Fuller was the first to describe the bicycle wheel in terms of tension and compression(Fuller) while Levin used it as an analogy to explain some of the complexities of the human body in terms of biotensegrity.Levin Here the outer rim and central hub are considered as compression elements held in place by a network of wire spokes in reciprocal tension. This type of wheel is a self-contained entity maintained in perfect balance throughout with no bending moments or torque, no fulcrum of action and no levers. He proposed that the scapula functions as the hub of such a wheel, in effect as a sesamoid bone, and transfers its load to the axial skeleton through muscular and fascial attachments. The sterno-clavicular joint is not really in a position to accept much compressional load and the transfer of axial compression across the gleno-humeral joint is at maximum only when loaded at 90o abduction.
Fuller was the first to describe the bicycle wheel in terms of tension and compression(Fuller) while Levin used it as an analogy to explain some of the complexities of the human body in terms of biotensegrity.Levin Here the outer rim and central hub are considered as compression elements held in place by a network of wire spokes in reciprocal tension. This type of wheel is a self-contained entity maintained in perfect balance throughout with no bending moments or torque, no fulcrum of action and no levers. He proposed that the scapula functions as the hub of such a wheel, in effect as a sesamoid bone, and transfers its load to the axial skeleton through muscular and fascial attachments. The sterno-clavicular joint is not really in a position to accept much compressional load and the transfer of axial compression across the gleno-humeral joint is at maximum only when loaded at 90o abduction.
The joint is essentially a frictionless inclined plane which means that it must rely heavily on ligamentous and muscular tension in all other positions. The humerus as a hub model would function equally well with the arm in any position. Interestingly, different parts of the gleno-humeral capsule that transfer specific tensional stresses can only do so if the capsule is intact, even if those stresses do not apparently pass through the missing parts,Moore but this makes perfect sense if the capsule is considered as a tensegrity sheet at a microscopic level.
In a similar way, the ulna can be compared to a hub within the distal humeral ‘rim’ of muscle attachments, where load bearing across the joint is significantly tensional, and compressional forces are contained within the bones and soft-tissue compartments, thus allowing the hand to lift much larger loads than would otherwise be the case (see the elbow page).
 The pelvis can also be compared with the wheel with the iliac crests, anterior spines, pubis and ischia representing the outer rim and the sacrum representing the hub tied in with strong sacro-iliac, sacro-tuberous and sacro-spinous ligaments. Similarly the femoral heads act like hubs within the ‘spokes’ of the ilio-femoral, pubo-femoral and ischio-femoral ligaments.
The pelvis can also be compared with the wheel with the iliac crests, anterior spines, pubis and ischia representing the outer rim and the sacrum representing the hub tied in with strong sacro-iliac, sacro-tuberous and sacro-spinous ligaments. Similarly the femoral heads act like hubs within the ‘spokes’ of the ilio-femoral, pubo-femoral and ischio-femoral ligaments.
All these examples are huge simplifications of the anatomy but they do indicate the potential of looking at the body in this different way. Standard descriptions of synovial joints as hinges is very misleading because most bones follow complex helical pathways during movement, and it has been shown in the knee joint that there is no continuous compression between bones and cartilage, even when they are pushed together.Hakkak The above biotensegrity description of the fascia as a complex system of tensioned fibres enclosing a multi-scale heterarchy of compressed compartments now offers an alternative means to explain this non-bone compression/joint spacing (see the elbow example).

 According to Wolff’s law, tensional forces remodel the bony contours and alter the positions and orientations of their attachments, contributing to the complexity of shapes apparent in the skeleton. As part of a biotensegrity structure, each attachment would influence all the others and distribute forces throughout the system with points of potential weakness avoided, which is in contrast to a solid rod or truss that is vulnerable to buckling. Such a mechanism would be an advantage in long-necked animals such as giraffes, camels and dinosaurs, where the load from the head is distributed throughout the neck, as opposed to a stress-ridden cantilever system such as the Forth Bridge.
According to Wolff’s law, tensional forces remodel the bony contours and alter the positions and orientations of their attachments, contributing to the complexity of shapes apparent in the skeleton. As part of a biotensegrity structure, each attachment would influence all the others and distribute forces throughout the system with points of potential weakness avoided, which is in contrast to a solid rod or truss that is vulnerable to buckling. Such a mechanism would be an advantage in long-necked animals such as giraffes, camels and dinosaurs, where the load from the head is distributed throughout the neck, as opposed to a stress-ridden cantilever system such as the Forth Bridge.

 The erect spine and bipedal weight bearing capability of humans has traditionally been viewed as a tower of bricks and compressed disc joints that transfer the body weight down through each segment until it reaches the sacrum; but a vertical spine is a relatively rarity amongst vertebrates. Most other species have little or no use for a compressive vertebral column which is frequently portrayed as a horizontal truss and cantilever support system. As the main difference in vertebrate anatomies is in the detail it seems reasonable to suppose that they have some structural properties in common. Tensegrities are omni-directional ie. they are stable irrespective of the direction of loading, and the spine, pelvis and shoulder all demonstrate this property (within physiological limits), enabling dancers to tip-toe on one leg and acrobats to balance on one hand.Levin
The erect spine and bipedal weight bearing capability of humans has traditionally been viewed as a tower of bricks and compressed disc joints that transfer the body weight down through each segment until it reaches the sacrum; but a vertical spine is a relatively rarity amongst vertebrates. Most other species have little or no use for a compressive vertebral column which is frequently portrayed as a horizontal truss and cantilever support system. As the main difference in vertebrate anatomies is in the detail it seems reasonable to suppose that they have some structural properties in common. Tensegrities are omni-directional ie. they are stable irrespective of the direction of loading, and the spine, pelvis and shoulder all demonstrate this property (within physiological limits), enabling dancers to tip-toe on one leg and acrobats to balance on one hand.Levin
RESPIRATORY SYSTEM OF THE BIRD
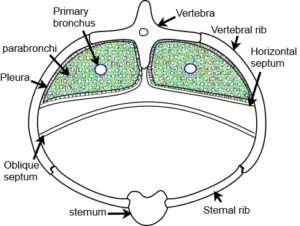 The respiratory system of the bird differs substantially from the mammalian lung; it is an exceptionally efficient gas exchanger that processes the large amounts of oxygen required to sustain flight. Some of the reasons for this are considered to be its geodesic design and heterarchical biotensegrity arrangement that mechanically couples each part into a functionally unified structure.Maina The volume of the bird lung is about 27% less than that of a mammal of similar body mass although the respiratory surface area is about 15% greater. The lung is attached to a rigid ribcage and its volume changes relatively little during a respiratory cycle (1.4%); instead, separate air sacs act like bellows and cause unidirectional and continuous ventilation. The air passages of the lung have a heterarchical arrangement with two-thirds of the lung volume taken up with several hundred parabronchi; their polygonal atrial openings each give rise to several funnel shape ducts (infundibulae) that terminate in numerous air capillaries, the terminal respiratory units (fig. ?). Both blood and air capillaries anastomose and interdigitate to form a tightly packaged three-dimensional network.
The respiratory system of the bird differs substantially from the mammalian lung; it is an exceptionally efficient gas exchanger that processes the large amounts of oxygen required to sustain flight. Some of the reasons for this are considered to be its geodesic design and heterarchical biotensegrity arrangement that mechanically couples each part into a functionally unified structure.Maina The volume of the bird lung is about 27% less than that of a mammal of similar body mass although the respiratory surface area is about 15% greater. The lung is attached to a rigid ribcage and its volume changes relatively little during a respiratory cycle (1.4%); instead, separate air sacs act like bellows and cause unidirectional and continuous ventilation. The air passages of the lung have a heterarchical arrangement with two-thirds of the lung volume taken up with several hundred parabronchi; their polygonal atrial openings each give rise to several funnel shape ducts (infundibulae) that terminate in numerous air capillaries, the terminal respiratory units (fig. ?). Both blood and air capillaries anastomose and interdigitate to form a tightly packaged three-dimensional network.
The parabronchi develop from epithelial cells that are compressed due to space restraint and naturally form hexagons with lumens that enlarge during development. This geodesic packing arrangement persists into the adult and makes the most economical utilization of space, thus maximizing the potential respiratory surface area. The constitutive parts of the parabronchus act together to function as an integrated unit that prevents the air capillaries from collapsing under compression and blood capillaries from distending with over-perfusion; mechanically, it is rather similar to the tensegrity bicycle wheel described above.Maina2010
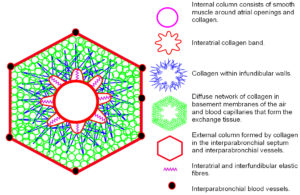 Intertwined smooth muscle bundles and collagenous tissue surround the atrial openings into the central lumen and form a complex helical arrangement. The collagen forms an intricate system of longitudinal, transverse and oblique fibres that connect to elastic fibres in the interatrial septa and floor of each atrium, and continue as the interfundibula septa that eventually becomes the basement membrane surrounding the exchange tissues. The smooth muscle, collagen and elastic fibres surrounding the atrial openings form an internal parabronchial column that lies next to the lumen. The collagenous septa and exchange tissues are also continuous with the interparabronchial septa that enclose the walls of larger blood vessels and form an external parabronchial column. The exchange tissues and associated septa are thus suspended between the internal and external parabronchial columns like the spokes in a bicycle wheel.
Intertwined smooth muscle bundles and collagenous tissue surround the atrial openings into the central lumen and form a complex helical arrangement. The collagen forms an intricate system of longitudinal, transverse and oblique fibres that connect to elastic fibres in the interatrial septa and floor of each atrium, and continue as the interfundibula septa that eventually becomes the basement membrane surrounding the exchange tissues. The smooth muscle, collagen and elastic fibres surrounding the atrial openings form an internal parabronchial column that lies next to the lumen. The collagenous septa and exchange tissues are also continuous with the interparabronchial septa that enclose the walls of larger blood vessels and form an external parabronchial column. The exchange tissues and associated septa are thus suspended between the internal and external parabronchial columns like the spokes in a bicycle wheel.
Contraction of smooth muscles around the atrium tenses the interatrial and interfundibula septae and stretches the elastic fibres, with collagen limiting their stretchability; the elastic fibres then act as energy-storage elements and recoil when the muscles relax. The interatrial, interfundibula and interparabronchial septa thus balance the centripetal force produced by contraction of the smooth muscle. An outward centrifugal force is also produced, by surface tension generated within the air capillaries and the prevailing intramural pressure in the interparabronchial arteries, and this is balanced by the elastic and inflexible collagen fibres. The parabronchus thus exists in a dynamically tensed state, with the inward pull of the atrial smooth muscles (internal column/wheel hub) ultimately counterbalanced by the interparabronchial septa (external column/wheel rim) and surrounding parabronchi. The morphology of the parabronchus and its constitutive parts thus fits every definition of a tensegrity structure.
MAMMALIAN LUNG ALVEOLI
The matrix surrounding alveoli is considered to be a biotensegrity structure. “The septa between alveoli are very thin and contain a single dense capillary network. They are supported by a fine network of fibres that are interwoven with the capillaries and anchored at both ends in axial fibres that form the network of alveolar entrance rings in the wall of alveolar ducts; and peripheral fibres that extend through interlobular septa towards the pleura. This allows the spreading of the capillaries by mechanical tension on the fibres. Because of this disposition of capillaries and fibres, alveoli in the mature lung are not structural units that can be separated: each of their walls is shared by two adjoining alveoli, both in terms of gas exchange with the capillary and with respect to mechanical support. Even the epithelial lining is shared by two adjacent alveoli as it extends through the pores of Kohn… This disposition of the fibre system makes the lung a tensegrity structure, which means that, in terms of mechanics, the integrity of lung parenchymal structure is exclusively ensured by the tension of the fibre continuum that supports alveolar walls and their capillaries. If one fibre is cut, this causes collapse of the septum followed by rearrangement of the adjacent parts, as occurs in emphysema.Weibel
Well, this page is the start of a vast amount of information that is accumulating about the value of biotensegrity to understanding biology, and the human body in particular. Because every part is in an inseparable relationship with every other part, no amount of writing can do justice to this subject on its own. So absorb what you can, read the books and it will transform the way the way we think about life and the treatment of its problems.

 Whenever nature uses the same principle in a variety of different situations there is probably an underlying energetic advantage to its appearance, and biological development and evolution will always favour those patterns and shapes that are the most energy-efficient in terms of stability, materials and mass. Even though they can appear to be rather complicated, at the most basic level, all living structures are the result of interactions between atomic forces and the orderly arrangements that they settle into.
Whenever nature uses the same principle in a variety of different situations there is probably an underlying energetic advantage to its appearance, and biological development and evolution will always favour those patterns and shapes that are the most energy-efficient in terms of stability, materials and mass. Even though they can appear to be rather complicated, at the most basic level, all living structures are the result of interactions between atomic forces and the orderly arrangements that they settle into. Biotensegrity recognizes that the forces of attraction and repulsion at the molecular scale are comparable with those of tension and compression at higher size scales, and are easily modelled using cables and struts, respectively. It is a simple re-evaluation of anatomy as a network of structures under tension and others that are compressed; parts that pull things together and others that keep them apart; basic physics!
Biotensegrity recognizes that the forces of attraction and repulsion at the molecular scale are comparable with those of tension and compression at higher size scales, and are easily modelled using cables and struts, respectively. It is a simple re-evaluation of anatomy as a network of structures under tension and others that are compressed; parts that pull things together and others that keep them apart; basic physics! The term ‘tensegrity’ is a combination of the words tension and integrity, and this structural system was first recognized as a new structural principle in 1948 by Kenneth Snelson (1927-2016), a young sculptor who produced many impressive works that he described as “…unveil[ing] the exquisite beauty of structure itself”. Tensegrity models are particularly interesting because the struts remain isolated and do not compress each other at any point because they are suspended within the tension network. The architect Buckminster Fuller (1895-1983) recognized them as part of his theory of Synergetics, the study of nature’s coordinate system that considers that all natural structures are inherent displays of the forces within them; and the cell biologist, Donald Ingber, has described the structural lattice (cytoskeleton) within cells as a tensegrity structure that regulates cell function and applies at every size scale in the body. Mechanical engineers also appreciate the distinctive properties of tensegrity structures and are producing robots for use in the exploration of space etc. Both biologists and engineers now recognize that the simple principles of tensegrity can be applied to understanding the behaviour of more complex structures, but because certain aspects are not transferable between these different fields, Stephen Levin introduced the term biotensegrity to distinguish between them.
The term ‘tensegrity’ is a combination of the words tension and integrity, and this structural system was first recognized as a new structural principle in 1948 by Kenneth Snelson (1927-2016), a young sculptor who produced many impressive works that he described as “…unveil[ing] the exquisite beauty of structure itself”. Tensegrity models are particularly interesting because the struts remain isolated and do not compress each other at any point because they are suspended within the tension network. The architect Buckminster Fuller (1895-1983) recognized them as part of his theory of Synergetics, the study of nature’s coordinate system that considers that all natural structures are inherent displays of the forces within them; and the cell biologist, Donald Ingber, has described the structural lattice (cytoskeleton) within cells as a tensegrity structure that regulates cell function and applies at every size scale in the body. Mechanical engineers also appreciate the distinctive properties of tensegrity structures and are producing robots for use in the exploration of space etc. Both biologists and engineers now recognize that the simple principles of tensegrity can be applied to understanding the behaviour of more complex structures, but because certain aspects are not transferable between these different fields, Stephen Levin introduced the term biotensegrity to distinguish between them.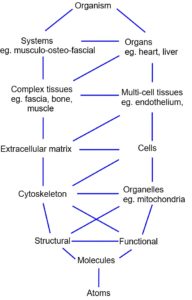 CONTENTS: Structural heterarchies; Cellular cytoskeleton and morphogenesis; Cell cortex; Helix; Collagen; Fascial system; Cranial vault; Spider silk; Shoulder joint, Elbow and pelvis; Respiratory system of the bird; Mammalian lung.
CONTENTS: Structural heterarchies; Cellular cytoskeleton and morphogenesis; Cell cortex; Helix; Collagen; Fascial system; Cranial vault; Spider silk; Shoulder joint, Elbow and pelvis; Respiratory system of the bird; Mammalian lung. Tensegrity heterarchies display a significant reduction in mass (with the ‘spaces’ filled with other smaller structures), and as tension always tries to reduce itself to a minimum, they automatically balance in the most energy-efficient configuration. Because every part influences every other part, potentially damaging forces can be distributed throughout the network and harmful stress concentrations avoided.
Tensegrity heterarchies display a significant reduction in mass (with the ‘spaces’ filled with other smaller structures), and as tension always tries to reduce itself to a minimum, they automatically balance in the most energy-efficient configuration. Because every part influences every other part, potentially damaging forces can be distributed throughout the network and harmful stress concentrations avoided.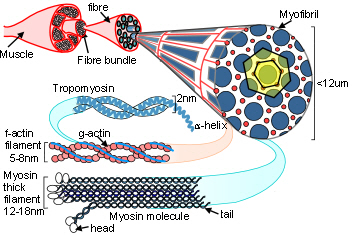
 CELLULAR CYTOSKELETON AND MORPHOGENESIS
CELLULAR CYTOSKELETON AND MORPHOGENESIS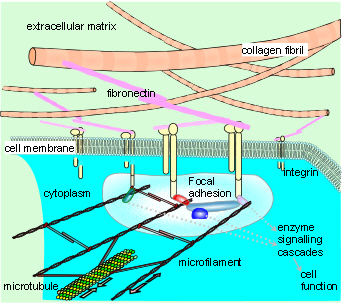 Experiments that allowed individual cells to assume certain shapes showed that those able to distort or spread had the highest rates of growth; rounded cells became apoptotic (died) while those intermediate in shape became quiescent and differentiated. Cells also tend to extend new motile processes (lamellipodia and filopodia) on sharp corners rather than blunt ones and this is linked with the cytoskeleton.
Experiments that allowed individual cells to assume certain shapes showed that those able to distort or spread had the highest rates of growth; rounded cells became apoptotic (died) while those intermediate in shape became quiescent and differentiated. Cells also tend to extend new motile processes (lamellipodia and filopodia) on sharp corners rather than blunt ones and this is linked with the cytoskeleton.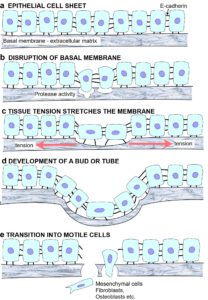 The transfer of tension from the ECM stimulates actomyosin tension generation, causing an increase in integrin binding and clustering, and the recruitment of more focal adhesion proteins that balance the ECM tension.
The transfer of tension from the ECM stimulates actomyosin tension generation, causing an increase in integrin binding and clustering, and the recruitment of more focal adhesion proteins that balance the ECM tension.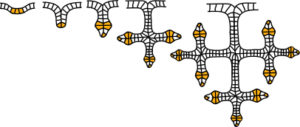 Branching can create a pattern similar to the ‘Koch snowflake’ fractal, and it has been found that the position of coronary artery lesions around the heart follows a pattern related to the Fibonacci sequence and Golden Mean, thus maximising perfusion of the myocardial bed.
Branching can create a pattern similar to the ‘Koch snowflake’ fractal, and it has been found that the position of coronary artery lesions around the heart follows a pattern related to the Fibonacci sequence and Golden Mean, thus maximising perfusion of the myocardial bed.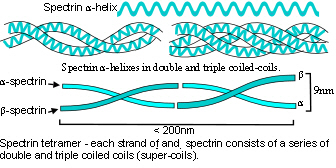
 CELL CORTEX
CELL CORTEX